Лес – сложная природная система, характерной чертой которой является динамичность. Природа леса многообразна и разные по характеру леса могут иметь различное назначение. Сложная природа леса стала доступна человеческому пониманию только в конце XIX в.
Лесоведение, как учение о лесе, о его природе, сформировалось в начале XX в. Основоположником лесоведения в России является проф. Г.Ф. Морозов, написавший фундаментальный труд «Учение о лесе» [3 — 4]. Он блестяще показал, что лес представляет собой явление сложное, где все части, компоненты взаимодействуют между собой и окружающей средой. Каждый компонент в отдельности и лес, как их совокупность, изменяются непрерывно и постепенно.

И.С. Мелехов [2] считает лесоведение естественно-исторической частью лесоводства, раскрывающей природу леса и рассматривающей вопросы биологии и экологии леса в целом и составляющих его компонентов.
Обычно лес определяют как совокупность деревьев на определенной площади, или часть земной поверхности, покрытой множеством деревьев. Следовательно, множественность деревьев – это важный признак леса, но недостаточный. Нельзя назвать лесом множество деревьев, пространственно отдаленных друг от друга. Множество деревьев составит лес только при наличии определенных качественных и количественных признаков. Совокупность деревьев можно будет назвать лесом лишь в том случае, когда число деревьев на единице площади достигнет определённой величины и множество деревьев, как сообщество, вступит во взаимодействие со средой, изменит эту среду настолько существенно, что эти изменения отразятся на самих деревьях. Только в этом случае совокупность деревьев можно назвать лесом.
Для понимания сущности леса необходимо сравнить деревья одной и той же породы, растущие в лесу и одиночно в одних и тех же почвенно-климатических условиях. Также необходимо изучить взаимодействие тех и других деревьев с окружающей средой.

Рис. 1. Сосна, выросшая в насаждении
При таком сравнении заметно различие деревьев по их форме и размерам. Дерево, выросшее на открытом месте, имеет небольшую высоту, часто шаровидную форму кроны, толстую, трещиноватую кору, толстые сучья, сбежистый ствол со слабым очищением от сучьев. Дерево, выросшее в лесу, характеризуется высоким полнодревесным стволом, хорошо очищенным от сучьев, высоко поднятой часто пирамидальной и компактной кроной с тонкими ветвями.
Таким образом, и по стволу и по кроне деревья, растущие в лесу, существенно отличаются от деревьев, выросших на открытом пространстве. Эти различия нельзя объяснить различием породы, поскольку сравниваются деревья одной породы. Нельзя объяснить их и различием в возрасте, так как сравниваются деревья одного возраста. Эти различия нельзя объяснить и случайными причинами, например болезнями, повреждением деревьев энтомовредителями, так как эти различия не единичный случай, а массовое, закономерное явления.

Рис. 2. Сосна, выросшая на открытом месте
Различия в положении сравниваемых деревьев заключаются в том, что одно из них растёт в лесу, в окружении себе подобных деревьев, в лесной обстановке, в тени, в безветрии и т.д., а другое растёт изолированно, вне этой лесной обстановки.
Лесная обстановка, в частности теневой режим, приводит к тому, что деревья усиленно растут вверх, образуя более высокие и стройные стволы. При этом стволы деревьев становятся более полнодревесными и прямыми. Это объясняется тем, что лесные деревья защищены от ветра и у них нет необходимости развивать устойчивый короткий и сбежистый ствол, как у деревьев, выросших на открытом пространстве. Вследствие этого же в лесу кроны деревьев более узкие, чем у свободно стоящих деревьев.
Длительные наблюдения лесоводов показали, что у деревьев, выросших в насаждении, возраст возмужалости, то есть период начала плодоношения, наступает на 10-20 лет позже, чем у деревьев, растущих на свободе. Г.Ф. Морозов также отмечает, что в лесу резче заметна периодичность плодоношения, так называемые семенные годы, в отличие от деревьев простора.
По данным проф. В.Д. Огиевского, если деревья, выросшие в насаждении, неожиданно выставить на простор (например, при рубке), то они не сразу увеличивают плодоношение, а только через некоторое время. Например, сосна обыкновенная требует для этого около 4 лет. За этот период деревья как бы приспосабливаются к новым условиям.
Таким образом, можно утверждать, что взаимовлияние деревьев в лесу и на открытом месте не ограничивается только изменением внешнего вида и процессов роста деревьев, но отражается также на процессах размножения.
Из-за теневой обстановки в лесу нижние ветви у лесных деревьев постепенно отмирают и отваливаются, поэтому крона у них сосредоточивается в верхней части ствола. Процесс отмирания нижних ветвей у лесных деревьев называется естественным очищением ствола от сучьев.

Рис. 3. Процесс очищения ствола от сучьев в сосновом древостое
Влияя друг на друга и на занятую ими территорию, деревья создают новые, лесные условия для себя и других организмов, определенным образом влияют на окружающую среду. Следовательно, причиной различий между лесными и нелесными деревьями является наличие или отсутствие особой лесной среды.
В результате приведённого сравнения можно сделать заключение о том, что лес – это не простое, а тесно взаимодействующее с окружающей средой множество деревьев. Это взаимодействие как раз и проявляется в резких отличиях лесных деревьев от деревьев, выросших вне леса. Деревья в лесу не только разнообразно влияют друг на друга, и на занятую этими деревьями среду, но и формирует свою собственную среду.
Г.Ф. Морозов первым дал определение лесу как сложному комплексу, все компоненты которого взаимодействуют между собой и с окружающей средой. Он писал, что «…лес не есть простая совокупность древесных растений, а представляет собой сообщество, или такое соединение древесных растений, в котором они проявляют взаимное влияние друг на друга, порождая тем целый ряд новых явлений, которые не свойственны одиноко растущим растениям». Далее Г.Ф. Морозов подчёркивает, что лесом будет называться «только такое сочетание древесных растений, которое характеризуется, во-первых, устойчивостью, во-вторых, взаимодействием друг на друга и, в-третьих, влиянием на занятую среду».
Н.М. Горшенин и А.И. Швиденко [1] характеризуют лес как элемент географического ландшафта, состоящий из совокупности древесных, кустарниковых, травянистых растений, животных и микроорганизмов, в своем развитии биологически взаимосвязанных, влияющих друг на друга и на внешнюю среду.
И.С. Мелехов [2] рассматривает лес, как сложный природный комплекс, занимающий определенную территорию, состоящий не только из деревьев, но и других компонентов растительного, животного и другого происхождения, биологически связанных со средой и взаимным влиянием друг на друга. Взаимоотношения между компонентами леса чрезвычайно многообразны и сложны. По выражению В.Н. Сукачева [7], «жизнь леса есть беспрерывная борьба за существование».
Г.Ф. Морозов отмечает, что взаимное влияние деревьев в лесу распространяется также и на внутреннее строение древесины деревьев. Это проявляется в изменении плотности, состава и анатомического строения древесины в пределах одной и той же породы, растущей на одной и той же почве, в одном и том же климате и зависит лишь от того, выросло ли дерево на свободе или в насаждении. Практика лесного хозяйства показывает, что хвойные деревья, выросшие на свободе, отличаются менее плотной древесиной по сравнению с древесиной деревьев, выросших в сомкнутых насаждениях.
В лесу постоянно и непрерывно протекают естественные процессы, которые обеспечивают его рост, развитие, формирование, отмирание и новое возобновление. Следовательно, лес является саморегулирующейся и самовозобновляющейся экологической системой.
Одним из наиболее важных и характерных естественных процессов в лесу является процесс изреживания деревьев с возрастом, что является закономерным результатом борьбы за существование, индивидуальной изменчивости, наследственности и естественного отбора. Процесс естественного изреживания заключается в уменьшении число деревьев на единице площади с возрастом. Этот процесс присущ древостоям всех пород как естественного, так и искусственного происхождения.
В процессе естественного изреживания значительная часть деревьев отмирает в силу физиологических причин — из-за дефицита воздушного и почвенного питания. Отпад деревьев в естественном лесу довольно значителен. На протяжении жизни древесного растения в результате процесса естественного изреживания отмирает более 95% деревьев. Если же исчислять отпад с момента появления всходов, то эта цифра увеличиться до 99,9% и даже более. Таким образом, выживает лишь одно из 10 тысяч древесных растений [2]. По меткому выражению проф. В.Г. Нестерова [5] в лесу царит исключительная смертность: вымирает до 95% и более деревьев.
Изреживание древостоев изменяется в зависимости от породы деревьев, их возраста и условий местопроизрастания. Древостои берёзы, осины, лиственницы, сосны изреживаются быстро, интенсивно и постепенно становятся сравнительно редкими. В то же время древостои таких пород, как бук, ель, пихта изреживаются медленно и остаются густыми. Например, в возрасте 40-50 лет в средних условиях число стволов на 1 га и площадь питания одного дерева у разных пород достигает примерно таких величин [5].
Таблица 1. Число деревьев на 1 га и площадь питания одного дерева
| № п/п | Порода | Число стволов
на 1 га |
Площадь
питания, м2 |
| 1. | Лиственница | 1000 | 10 |
| 2. | Берёза | 1250 | 8 |
| 3. | Дуб | 1500 | 7 |
| 4. | Осина | 1550 | 6 |
| 5. | Сосна | 1250 | 5,5 |
| 6. | Ель | 3250 | 3 |
| 7. | Пихта | 5000 | 2 |
Различия между числом деревьев на единице площади и площадью питания деревьев объясняется различной потребностью древесных пород в жизненном пространстве, как в почве, так и над почвой.
Процесс естественного изреживания древостоев идет с различной интенсивностью на протяжении жизни леса. Вначале, когда молодые растения менее стойки в борьбе за существование, изреживание древостоев идёт наиболее интенсивно. Когда же лес становится спелым, процесс изреживания постепенно затухает.
Наглядное представление об отпаде деревьев с возрастом дают данные А.В. Тюрина по сосновым лесам Архангельской области:
Таблица 2. Отпад деревьев сосны с возрастом
| Возраст, лет | 20 | 40 | 60 | 80 | 100 | 120 | 140 | 160 |
| Число деревьев, шт/га | 7000 | 2525 | 1482 | 970 | 755 | 590 | 522 | 467 |
Следовательно, отпад деревьев происходит с неодинаковой интенсивностью на разных этапах жизни леса. Наиболее острая борьба за существование происходит в молодом возрасте, примерно до 40- 60 лет, о чём свидетельствует наибольший отпад деревьев в этом возрасте. По данным И.С. Мелехова, наибольшая интенсивность отпада в сосновом лесу наблюдается примерно в 40 лет (около 80%), на Севере – в 60 лет.
В каждом возрасте лес имеет свою густоту стояния деревьев на единице площади. В начале жизни древесных растений в лесу (в стадии всходов) на 1 га площади может насчитываться 400-500 тыс. растений, в молодняках – около 10 тыс. штук, в спелом лесу их остается всего около 500 деревьев. Это нормальная густота стояния деревьев, без которой не может сформироваться лес.
Густота стояния деревьев на единице площади связана с площадью питания одного дерева. По данным В.Г. Нестерова [5], в средних условиях (III бонитет) одно сосновое дерево имеет следующую площадь питания:
Таблица 3. Площадь питания деревьев сосны по возрастным группам
| В. | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 | 100 | 110 | 120 | 130 | 140 |
| П | 1,6 | 2,7 | 4,3 | 5,5 | 7,7 | 9,1 | 12,5 | 14,3 | 16,6 | 18,2 | 20,0 | 22,2 | 25 |
Примечание: В – возраст деревьев, лет; П – площадь питания дерева, м2.
Таким образом, площадь питания одного дерева за период от 20 до 140 лет возросла с 1,6 до 25 м2, то есть в 16 раз.
Различна также интенсивность изреживания древостоев в разных лесорастительных и почвенно-климатических условиях. По мере ухудшения лесорастительных условий количество деревьев одного и того же возраста сохранившихся в процессе изреживания на 1 га увеличивается. Так же увеличивается число деревьев на 1 га на бедных почвах и в худшем климате.
Взаимосвязь и взаимовлияния деревьев одной породы в лесу в условиях нормальной естественной густоты стояния можно рассматривать как связь частей целого сообщества. Они создают друг другу различную среду, необходимую как для каждого дерева в отдельности, так и для всего сообщества в целом. Деревья теневого режима, умеренных температур, малой потребности во влаге и пище находятся под пологом других деревьев, которые образуют верхний полог и по своей природе требуют больше света и своими мощными корнями добывают из почвы большие количества пищи и влаги.
Следовательно, можно констатировать следующее: благодаря взаимному влиянию древесных растений друг на друга различия в наследственных и индивидуальных биоэкологических особенностях деревьев в лесу приводит их к острому соперничеству, то есть к жёсткой борьбе за существование.
Для одновозрастного древостоя характерна дифференциация деревьев по росту и развитию: среди одновозрастных деревьев можно выделить очень крупные, средние и очень мелкие деревья с различной степенью жизнеспособности. Деревья в лесу разнообразны, в то же время они обязательно несут признаки быстрого или замедленного развития, которые часто называют признаками стадийной молодости или старости. В.Г. Нестеров предлагает называть их признаками физической молодости и старости. Признаки дифференциации деревьев проявляются в изменчивости и форме деревьев, в сроках и обилии плодоношения, в качестве древесины, в морфолого-таксационных и физиологических показателях деревьев, в уровне их жизнеспособности и заболеваемости. Исследований заболеваний деревьев в лесу в зависимости от принадлежности их к различным классам роста и развития недостаточно. В этом плане представляет интерес исследования Ю.В. Синадского по зависимости распространения сосновой губки от классов роста и развития деревьев в сосняках Бузулукского бора и Б.П. Чуракова [8] – по степени зараженности деревьев разных классов роста смоляным раком, сосновой и корневой губками в ленточных борах Алтайского края.
Дифференциация деревьев по размерам в лесу даже при одном возрасте настолько велика, что одни особи могут быть мельче других по объёму в 10-15 раз [6].
По данным А.В. Тюрина, все леса характеризуются единой общей закономерностью строения, согласно которой во всяком одновозрастном древостое, если высоту среднего дерева принять за 1, то самое низкое дерево будет иметь высоту 0,80, а самое высокое – 1,15. Диаметр деревьев изменяется ещё сильнее. Если средний диаметр дерева в любом однородном участке леса принять за 1, то самое тонкое дерево будет иметь диаметр 0,5, а самое толстое – 1,7. Ещё более заметная разница наблюдается в объёмах деревьев. Во всяком древостое самое маленькое дерево составляет по объёму 0,221 долю от среднего, а самое большое превышает среднее в 2,991 раза.
Дифференциация деревьев в лесу по размерам издавна привлекала внимание лесоводов. Они стремились установить её размах, изучить закономерности изменчивости деревьев, познать роль дифференциации в жизни леса, раскрыть причины её, научиться управлять этим процессом и использовать его в лесохозяйственной практике.
Основными причинами дифференциации деревьев в лесу являются наследственность, индивидуальная изменчивость и выживаемость, связанная с неоднородными условиями среды, в которую попадают семена, В лесоводстве в целях ухода за лесом применяются различные классификации деревьев по степени их дифференциации – развитости.
Стали появляться различные классификации деревьев, и их было предложено огромное количество. В.Г. Нестеров в своей монографии «Вопросы современного лесоводства» приводит характеристики 30 наиболее интересных классификаций деревьев. Все появившиеся в истории лесоводства классификации он делит на 3 категории:
- первоначальные, или старые, когда деревья в лесу подразделялись по рангам господства и угнетённости, либо просто по размерам (8);
- усовершенствованные, или модернизированные, в которых деревья делились ещё на разные категории в зависимости от технических качеств (18);
- новые классификации, подразделяющие деревья, как по росту, так и по развитию – биологическому качеству (4).
Первоначально лесоводы стали выделять в лесу господствующие и угнетённые деревья. К первым относили крупные и хорошо развитые деревья; ко вторым – мелкие, недоразвитые, затенённые и угнетённые.
Позднее делались попытки более детальных подразделений деревьев. Например, известный немецкий лесовод Котта в 1844 г. подразделял деревья на 4 класса:
- господствующие;
- подчинённые;
- угнетённые;
- мёртвые.
В 1848 г. немецкий лесовод Буркгард предложил разделить деревья на 6 классов в зависимости от их высоты, положения и развития кроны. Но наибольшее распространение среди лесоводов получила классификация деревьев по росту другого немецкого лесовода Крафта, разработавшего её в 1884 г. Эта классификация имеет много общего классификацией Буркгарда.
Крафт выделил пять классов, обозначив их римскими цифрами: I – исключительно крупные, господствующие мощные деревья с толстыми стволами и сильно развитыми кронами (5%), II – крупные, господствующие деревья с хорошо развитыми стволами и большими кронами (30-40%), III – средние деревья (согосподствующие, по Морозову, кандидаты на угнетение), характеризующиеся меньшей высотой и диаметром ствола, более узкой кроной (20-40%), IV – угнетённые, мелкие, тонкие, отстающие в росте деревья с узкими и слабыми кронами (10-20%). Среди них выделяются две категории деревьев: IVа – равномерно угнетённые со всех сторон и имеющие узкие расходящиеся во все стороны кроны; IVб – флагообразные, с кроной развитой лишь в одну сторону; V – отмирающие (Vа) и мертвые (Vб) деревья (до10%).
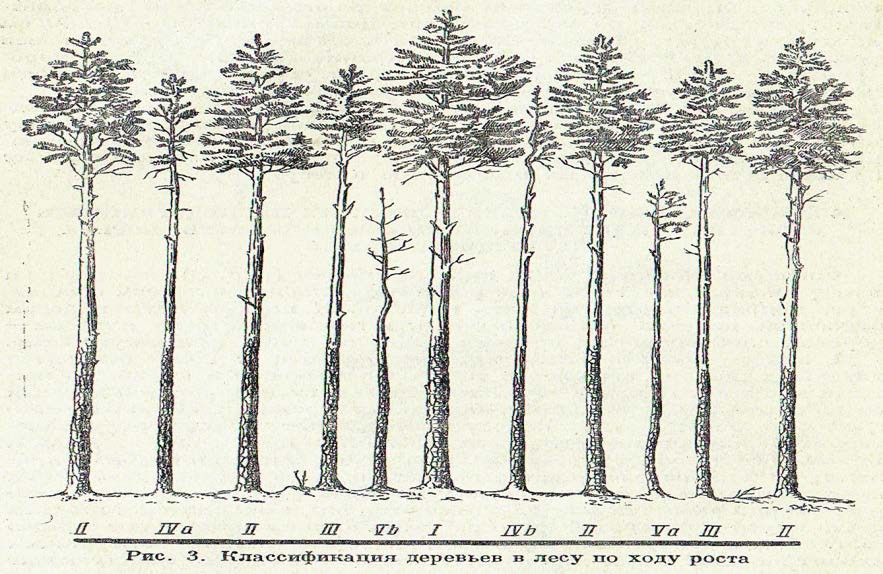
Рис. 4. Классификация деревьев в лесу по росту (Крафта)
Изучение лесоводами плодоношения деревьев, относящихся к разным классам господства и угнетения, показало, что урожайность таких деревьев очень сильно отличается. Например, исследования А.Н. Соболева и А.Ф. Фомичёва в лесах Ленинградской области показали, что в хороший семенной год возмужалый еловый лес дает следующие величины урожая по классам Крафта: деревья I класса – 100%, II – 88%, III – 37%, IV – 0,5%, V – 0%.
Среди отечественных классификаций по аналогичным Крафту принципам следует отметить подразделение деревьев, предложенное в 1915 г. учёным лесоводом В.И. Переходом. Он классифицировал деревья в сомкнутом древостое на 3 группы:
- господствующие деревья, то есть развивающиеся сильнее остальных и потому играющие первостепенную роль;
- индифферентные, то есть не влияющие на рост других деревьев и сами не находящиеся под влиянием;
- угнетённые деревья, отстающие от соседних в росте и притенённые более сильными, господствующими деревьями.
Старые (первоначальные) классификации деревьев постепенно стали подвергаться критике. Критиковали их главным образом за недостаточность подразделения деревьев только по принципу господства и угнетённости, указывали на необходимость деления деревьев ещё и по техническим качествам.
Примером усовершенствованной классификации деревьев по техническим качествам стала классификация деревьев немецкого лесовода К.Гека, разработанная им в конце XIX в. Согласно этой классификации, деревья делятся на 7 классов:
- I – прямые длинные стволы;
- II – прямые деловые, но более короткие;
- III – кривые, суковатые деревья;
- IV – двойчатки;
- V – сильно корявые деревья;
- VI – порослевые деревья;
- VII – больные деревья.
В этой классификации деревья выделяются не только по принципу господства и угнетённости, но и по техническим качествам.
В 1947 г. отечественный лесовод И.П. Пряхин предложил перестроить классификацию деревьев по Крафту в классификацию деревьев «по жизнеспособности и выживанию». Им были предложены следующие ступени:
- Первая ступень выживания – деревья вполне жизнеспособные, здоровы, обгоняющие в росте большинство и выходящие из полога древостоя.
- Вторая ступень выживания – деревья жизнеспособные, здоровые, составляющие большинство и образующие верхний полог древостоя.
- Третья ступень – деревья переходные, здоровые, отстающие в росте от большинства, но способные к переходу во вторую ступень.
- Четвертая ступень – деревья условно переходные, часто больные, отстающие в росте от деревьев третьей ступени, но способные к переходу в эту ступень.
- Пятая ступень – деревья ненадёжные, часто больные, отстающие в росте от деревьев четвертой ступени явно сомнительные в смысле перехода их в эту ступень.
Из других классификаций деревьев необходимо отметить классификации М.Д. Данилова (1948), П.В.Воропанова (1950), Д.И. Дерябина (1953) и В.Г. Нестерова (1961). Наибольший интерес из перечисленных классификаций представляет классификация деревьев по росту и развитию В.Г. Нестерова. Он предложил подразделять деревья не только по росту, но и по степени из развития. В.Г. Нестеров отмечает, что «…рост и развитие – явления, хотя и тесно связанные, существующие в единстве, но всё же не представляют одного и того же и всегда находятся в некотором противоречии между собой». Далее он пишет «…рост, величина деревьев лучше всего определяются их высотой, а развитие – главным образом плодоношением, которое, в свою очередь связано с изменением прироста по высоте».
В своей классификации В.Г. Нестеров выделяет 3 класса деревьев по росту с подразделением их на 2 подкласса (а и б) по степени
развития. В общем виде классификация деревьев по росту и развитию выглядит следующим образом:
- класс – деревья быстрого роста: а – медленного развития, б – быстрого развития.
- класс – деревья замедленного роста: а – медленного развития, б – быстрого развития.
- класс – деревья слабого роста: а – не развивающиеся, б – сильно отставшие, отмирающие и усохшие, слабо развитые и не оставившие потомства.
Под медленно развивающимися автор подразумевает деревья любого размера, если они длительно растут по высоте и поздно вступают в фазу плодоношения, долго не стареют. К быстро развивающимся он относит деревья, которые в раннем возрасте заканчивают рост в высоту, скоро начинают плодоносить и быстро отмирают.
Процесс дифференциации деревьев зависит от породы, возраста и условий окружающей среды. У всех древесных пород на богатых и оптимально увлажненных почвах дифференциация деревьев выражены сильнее, чем на бедных и сухих почвах, где деревья растут медленнее.
В связи с переходом лесного хозяйства на селекционно- генетическую основу производится селекционная инвентаризация наиболее производительных насаждений, при которой все деревья классифицируются на следующие категории:
- Плюсовые деревья – это деревья, значительно превосходящие по комплексу хозяйственно-ценных признаков и свойств деревья того же возраста, растущие с ними в одинаковых условиях. При этом учитывают показатели не только роста и качества, но и устойчивость к вредителям, болезням, и неблагоприятным факторам среды, а также характер плодоношения. В чистых одновозрастных насаждениях плюсовые деревья должны, как правило, иметь превышение над средним диаметром насаждения на 60-70%, а по высоте – на 15%. При недостатке или отсутствии таких деревьев допускается отбор деревьев, превышающих диаметр насаждения не менее чем на 30%, а высоту – не менее чем на 10%.
- Нормальные деревья – это деревья, составляющие основную часть насаждения, хорошие и средние по силе роста, качеству и состоянию.
- Минусовые деревья – это все отстающие деревья, диаметр которых составляет 80% и менее от среднего диаметра насаждения, а также деревья большего диаметра, но кривые, суковатые, с сильными утолщениями на стволе, свилеватые и косослойные, с плохо развитой кроной, двойчатки, больные, поврежденные стволовыми вредителями, с сильными механическими повреждениями.
Таким образом, явления изреживания, очищения ствола от сучьев и дифференциации деревьев объясняются, во-первых, различными наследственными свойствами растений, во-вторых, их индивидуальной изменчивостью и, в третьих, естественным отбором.
Список литературы:
- Горшенин Н.М., Швиденко А.Н. Лесоводство. Львов, 1977.
- Мелехов И.С. Лесоведение. М.: Изд-во МГУЛ, 1999
- Морозов Г.Ф. Избранные труды. Т.1. М.: Изд-во «Лесная пром-сть», 1970
- Морозов Г.Ф. Избранные труды. Т.2. М.: Изд-во «Лесная пром-сть», 1971
- Нестеров В.Г. Общее лесоводство. М.: Сельхозгиз, 1960.
- Набатов Н.М. Лесоводство. М.: Изд-во МГУЛ, 1997
- Сукачёв В.Н., Дылис Н.В. Основы лесной биогеоценологии. М.: Изд-во «Наука», 1964
- Чураков Б.П. Взаимоотношения патогенных грибов с древесными растениями. М.: Изд-во МГУ им. М.В.Ломоносова, 1993.
